EPITHELIAL TISSUE
Despite its complexity, the human body is composed of only four basic types of tissue: EPITHELIAL, CONNECTIVE, MUSCULAR, and NERVOUS. These tissues, which are formed by cells and molecules of the extracellular matrix, exist not as isolated units but rather in association with one another and in variable proportions, forming different organs and systems of the body. Also of great functional importance are the FREE CELLS found in body fluids such as blood and lymph.

Most organs can be divided into two components: parenchyma, which is composed of the cells responsible for the main functions typical of the organ, and stroma (mesenchyma), which is the supporting tissue.
FUNCTIONS:
The principal functions of epithelial (Gr. epi, upon, + thele, nipple) tissues are the covering and lining of surfaces (eg, skin, intestines), absorption (eg, intestines), secretion (eg, glands), sensation (eg, gustative and olfactory neuroepithelium), and contractility (eg, myoepithelial cells). Because epithelial cells line all external and internal surfaces of the body, everything that enters or leaves the body must cross an epithelial sheet.
THE FORMS & CHARACTERISTICS OF EPITHELIAL CELLS:
The forms and dimensions of epithelial cells range from high columnar to cuboidal to low squamous cells.
Epithelial cell nuclei have distinctive shapes, varying from spherical to elongated or elliptic. The form of the nuclei of epithelial cells corresponds roughly to the cell shape; thus, cuboidal cells have spherical nuclei and squamous cells have flattened nuclei. The long axis of the nucleus is always parallel to the main axis of the cell.
Almost all epithelial cells, whether lining a surface or forming gland units, rest on a connective tissue. In the case of epithelia that line the cavity of internal organs (especially the digestive, respiratory, and urinary systems) this layer of connective tissue is often called lamina propria. The lamina propria not only serves to support the epithelium but also provides nutrition and binds it .
The portion of the epithelial cells that faces the connective tissue is called the basal pole, whereas the opposite side, usually facing a space, is called the apical pole. The surface of the apical pole is also called the free surface, whereas the surfaces that are apposed to neighbor cells are called lateral surfaces.
Basal Lamina & Basement Membrane
Most epithelial cells are separated from the connective tissue by a sheet of extracellular material called the basal lamina. This structure is visible only with the electron microscope, where it appears as a dense layer, 20–100 nm thick, consisting of a delicate network of very fine fibrils (lamina densa) .In addition, basal laminae may have an electron-lucent layer on one or both sides of the lamina densa, called lamina rara or lamina lucida. Between cell layers without intervening connective tissue, such as in lung alveoli and in the renal glomerulus ,the basal lamina is thicker as a result of fusion of the basal laminae of each epithelial cell layer.


The main components of basal laminae are type IV collagen, the glycoproteins laminin and entactin, and proteoglycans (eg, the heparan sulfate proteoglycan called perlecan). In some instances, reticular fibers are closely associated with the basal lamina, forming the reticular lamina. The term basement membrane is used to specify a periodic acid–Schiff (PAS)-positive layer, visible with the light microscope, present beneath some epithelia.

Intercellular Adhesion & Intercellular Junctions
Several membrane-associated structures contribute to cohesion and communication between cells. They are present in most tissues but are ,prominent in epithelium. Adhesion is due in part to the binding action of a family of transmembrane glycoproteins called cadherins.



Tight junctions, or zonulae occludens (singular, zonula occludens), are the most apical of the junctions. The Latin terminology gives important information about the geometry of the junction. "Zonula" refers to the fact that the junction forms a band completely encircling the cell , and "occludens" refers to the membrane fusions that close off the intercellular space.
In many epithelia, the next type of junction encountered is the zonula adherens. This junction encircles the cell and provides for the adhesion of one cell to its neighbor. A noteworthy feature of this junction is the insertion of numerous actin filaments into electron-dense plaques of material on the cytoplasmic surfaces of the junctional membranes. The filaments belong to the terminal web, a web of actin filaments, intermediate filaments, and spectrin found close to the free surface.
Gap or communicating junctions can occur almost anywhere along the lateral membranes of epithelial cells. The individual unit of the gap junction is called a connexon. Each connexon is formed by six gap junction proteins called connexins, which join together leaving a hydrophilic pore about 1.5 nm in diameter in the center. Connexons of adjacent cells are aligned to form a hydrophilic channel between the two cells . Each gap junction is formed by tens or hundreds of aligned pairs of connexons. A typical example is heart muscle cells, where gap junctions are, to a great degree, responsible for the heart's coordinated beat.
The final type of junction is the DESMOSOME (Gr. desmos, band, + soma, body), or MACULA ADHERENS. The desmosome is a complex disk-shaped structure at the surface of one cell that is matched with an identical structure at the surface of the adjacent cell. The cell membranes in this region are very straight and are frequently somewhat farther apart (>30 nm) than the usual 20 nm. On the cytosolic side of the membrane of each cell and separated from it by a short distance is a circular plaque of material called an ATTACHMENT PLAQUE, made up of at least 12 different proteins. In epithelial cells, groups of INTERMEDIATE CYTOKERATIN FILAMENTS are inserted into the attachment plaque or make hairpin turns and return to the cytoplasm. Because intermediate filaments of the cytoskeleton are very strong, desmosomes provide a firm adhesion among the cells. In nonepithelial cells, the intermediate filaments attached to desmosomes are made not of cytokeratin but of other proteins, such as desmin or vimentin. Proteins of the cadherin family participate in the adhesion provided by desmosomes
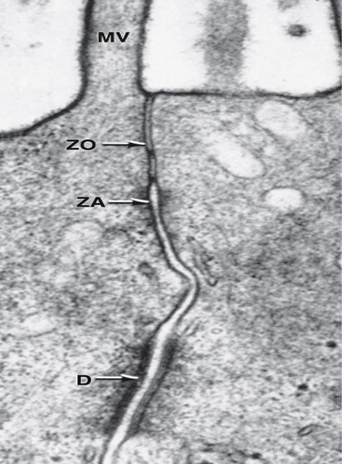





Specializations of the Cell Surface
Microvilli
When viewed in the electron microscope, the majority of cells from several different tissues are seen to have cytoplasmic projections. These projections, the microvilli (Gr. mikros, small, + L. villus, tuft of hair), are fingerlike extensions measuring about 1 m high and 0.08 m wide. They are found mainly on the free cell surface. Hundreds of microvilli are found in absorptive cells, such as the lining epithelium of the small intestine and the cells of the proximal renal tubule In these absorptive cells the glycocalyx is thicker than it is in most other cells. The complex of microvilli and glycocalyx may be seen with the light microscope and is called the brush, or striated, border.

As seen at higher magnification in micrograph, the cytoplasmic core of each microvillus contains actin microfilaments F which insert into the terminal web, a specialisation of the actin cytoskeleton lying immediately beneath the cell surface. At the periphery of the cell the terminal web is anchored to the zonula adherens .At the tip of the microvillus, the filaments attach to an electron-dense part of the plasma membrane. The microfilaments maintain stability of microvilli and may also mediate some contraction and elongation of the microvilli.


STEREOCILIA
Stereocilia are long, nonmotile extensions of cells of the epididymis and ductus deferens that are actually long and branched microvilli and should not be confused with true cilia. Stereocilia increase the cell surface area, facilitating the movement of molecules into and out of the cell.

CILIA & FLAGELLA
Cilia are cylindrical, motile structures on the surface of some epithelial cells, 5–10 m long and 0.2 m in diameter. They are surrounded by the cell membrane and contain a central pair of isolated microtubules surrounded by nine pairs of microtubules. The two microtubules of the peripheral pairs are joined to each other.

Cilia are inserted into BASAL BODIES, which are small cylindrical structures at the apical pole just below the cell membrane .Basal bodies have a structure analogous to that of the centrioles.
In living organisms, cilia have a rapid back-and-forth movement. Ciliary movement is frequently coordinated to permit a current of fluid or particulate matter to be propelled in one direction over the epithelial surface. Adenosine triphosphate (ATP) is the source of energy for ciliary motion. A ciliated cell of the TRACHEA is estimated to have about 250 cilia.
Flagella, present in the human body ONLY IN SPERMATOZOA, are similar in structure to cilia but are much longer and are limited to one flagellum per cell.








ALL BLOOD VESSELS ARE LINED WITH A SIMPLE SQUAMOUS EPITHELIUM CALLED ENDOTHELIUM

THE SIMPLE SQUAMOUS EPITHELIUM THAT COVERS THE BODY CAVITIES (THE ABDOMINAL CAVITY IN THIS CASE) IS CALLED MESOTHELIUM. PT STAIN. MEDIUM MAGNIFICATION

SIMPLE CUBOIDAL EPITHELIUM (ARROW) FROM KIDNEY COLLECTING TUBULES. PT STAIN. LOW MAGNIFICATION

SIMPLE COLUMNAR EPITHELIUM FORMED BY LONG CELLS WITH ELLIPTICAL NUCLEI.

STRATIFIED SQUAMOUS NONKERATINIZED (MOIST) EPITHELIUM OF THE ESOPHAGUS. THE MOST SUPERFICIAL CELLS (ARROW) HAVE THE FORM OF VERY THIN SCALES. PT STAIN. MEDIUM MAGNIFICATION.
STRATIFIED TRANSITIONAL EPITHELIUM OF THE URETHRA. THE RED-STAINED BASEMENT MEMBRANE BETWEEN THE EPITHELIUM AND THE UNDERLYING LOOSE CONNECTIVE TISSUE IS INDICATED BY ARROWS.

PSEUDOSTRATIFIED COLUMNAR EPITHELIUM OF THE TRACHEA, FORMED BY LONG AND SHORT CELLS. AS SOME CELLS DO NOT REACH THE SURFACE OF THE EPITHELIUM THEIR NUCLEI ARE PRESENT IN DIFFERENT HEIGHTS OF THE EPITHELIAL LAYER. MUCUS-SECRETING CELLS, CALLED GOBLET CELLS (ARROW), INTERMINGLE WITH CILIATED LINING CELLS.
GLANDULAR EPITHELIA
- Glandular epithelia are formed by cells specialized to produce secretion. The molecules to be secreted are generally stored in the cells in small membrane-bound vesicles called secretary granules.
- Glandular epithelial cells may synthesize, store, and secrete proteins (eg, pancreas), lipids (e.g., adrenal, sebaceous glands)
- complexes of carbohydrates and proteins (eg, salivary glands).
- The mammary glands secrete all three substances.
TYPES OF GLANDULAR EPITHELIA
The epithelia that form the glands of the body can be classified according to various criteria. Unicellular glands consist of isolated glandular cells, and multicellular glands are composed of clusters of cells. An example of a unicellular gland is the GOBLET CELL of the lining of the small intestine, or of the respiratory tract. The term "gland," however, is usually used to designate large, complex aggregates of glandular epithelial cells, such as in the salivary glands and the pancreas.

Section of large intestine showing goblet cells (arrows) secreting mucus to the extracellular space. The mucus precursor stored in the cytoplasm of the goblet cells is also stained in a dark color.

The nucleus of the goblet cell on the right is outside the plane of section, the nucleus N of the other being typically highly condensed showing a goblet cell in horizontal section). The cytoplasm is packed with rough endoplasmic reticulum rER; a few mitochondria M are present. A prominent Golgi apparatus G is found in the supranuclear region although it is barely visible at this magnification.
The protein component of mucigen is synthesised by the rough endoplasmic reticulum and passed to the Golgi apparatus where it is combined with carbohydrate and packaged into membrane-bound, secretory granules containing mucigen Mu

Formation of glands from covering epithelia. Epithelial cells proliferate and penetrate the connective tissue.

 Simple tubular glands, LARGE INTESTINE
Simple tubular glands, LARGE INTESTINE
Simple coiled tubular glands,
Sweat glands are almost the only example of simple coiled tubular glands.

Simple branched tubular glands
mainly in the stomach.

In this example of the mucus-secreting glands of the penile urethra, the secretory cells are pale stained compared to the non-secretory cells lining the urethra U. Note that the term acinus can be used to describe any rounded exocrine secretory unit.

Sebaceous glands provide a good example of simple branched acinar glands. Each gland consists of several secretory acini A that empty into a single excretory duct; the excretory duct E is formed by the stratified epithelium surrounding the hair shaft. The mode of secretion of sebaceous glands is HOLOCRINE, i.e. the secretory product, sebum, accumulates within the secretory cells and is discharged by degeneration of the cells.

Brunner's glands of the duodenum, as shown in this example, are described as compound branched tubular glands. Although difficult to visualise here, the duct system D is branched, thus defining the glands as compound glands and the secretory portions S have a tubular form, which is branched and coiled.

Compound acinar glands are those in which the secretory units are acinar in form and drain into a branched excretory ducts E system. The pancreas .


Compound tubulo-acinar glands have three types of secretory units; namely BRANCHED TUBULAR, BRANCHED ACINAR and branched tubular with acinar end-pieces called DEMILUNES. THE SUBMANDIBULAR SALIVARY GLAND shown here is the classic example. It contains two types of secretory cells, mucus-secreting cells and serous cells; the former are pale but the latter, which have a protein-rich secretion (digestive enzymes), stain strongly due to their large content of rough endoplasmic reticulum. Generally, the mucous cells form tubular components T whereas the serous cells form acinar components A and demilunes D. Part of an excretory duct E is also seen in the lower left corner of the micrograph.
Based on how the secretory products leave the cell, glands can be classified as merocrine (Gr. meros, part, + krinein) or holocrine (Gr. holos, whole, + krinein). In merocrine glands (eg, the pancreas), the secretory granules leave the cell by exocytosis with no loss of other cellular material. In holocrine glands (eg, sebaceous glands), the product of secretion is shed with the whole cell—a process that involves destruction of the secretion-filled cells. In an intermediate type—the apocrine (Gr. apo, away from, + krinein) gland—the secretory product is discharged together with parts of the apical cytoplasm,mammary gland
General Biology of Epithelial Tissues
POLARITY
In many types of epithelial cells the distribution of organelles and membrane proteins is different when comparing the basal and apical poles of the cell. This differential and stable organization of cell components is called polarity. This means that different parts of the cell may have different functions.
INNERVATION
Most epithelial tissues receive a rich supply of sensory nerve endings from nerve plexuses in the lamina propria. The exquisite sensitivity of the cornea, the epithelium covering the anterior surface of the eye, is due to the great number of sensory nerve fibers that ramify between corneal epithelial cells.
RENEWAL OF EPITHELIAL CELLS
Epithelial tissues are labile and their cells are renewed continuously by means of mitotic activity. The renewal rate is variable; it can be fast in tissues such as the intestinal epithelium, in which cells are replaced every week, or slow, as in the liver and the pancreas. In stratified and pseudostratified epithelial tissues, mitosis takes place within the germinal layer, closest to the basal lamina, which contains the stem cells.
0 comments:
Post a Comment